(gas-tro-pod-a: Latin meaning: gaster=stomach pous=foot :
stomach - foot!
in Greek: gastro (gastra = stomach) poda ( podia = feet, as suffix the format
is –poda where means having feet, "Podi" in single, "Podia"
in plural)
(Thank you to AFENTAKIS Andreas for adding the
Greek translations on this page)
|
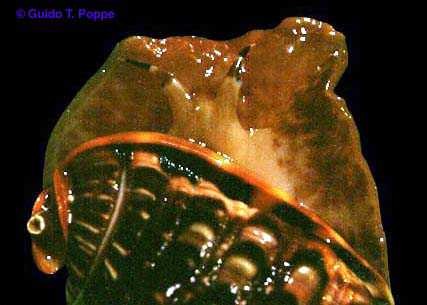
Cypraecassis
rufa
(by permission of Guido Poppe)
|
The
gastropoda is the largest and most certainly the
best-known Class of all the molluscs. They are the most successful
of the molluscan classes, and occupy almost every habitat on earth, from
desserts to high mountains, fields, forests, lakes, streams and oceans
- and most probably your back yard!! It is the only class to contain
species that have ventured permanently on to land. (To do this,
snails evolved an efficient gliding foot, eyes, an aggressive eating mechanism
and a pulmonary system for breathing.)
|
Gastropods
also inhabit every niche in the ocean from the intertidal zone to the deepest
ocean trenches. Over 15,000 fossil forms have been described and over 40,000
species exist today. They are, scientists theorize, now at the peak of
their evolutionary development.
Gastropods exhibit the least change from the ancestral molluscan plan of
all the molluscs. The pretorsion (pre = before, torsion = twisting) shell of
the ancestral spiraled gastropods resembled a coiled garden hose flat lying
on the ground. This plano-spiraled (i.e., coiled all in one plane - flat!)
shell was symmetrical. Having each coil lying outside the other was a
great disadvantage as it was not very compact nor was it easy to carry around
as the diameter could become very great (some fossil gastropods have been found
with shells measuring 8 feet (2.5m!!) in diameter - that's heavy-duty hauling!!!).
Then the gastropods underwent a very significant change in their evolutionary
path. This change was the twisting or �torsion � that the body
underwent. Most of the body located behind the head, including the visceral
mass, mantle and mantle cavity, was twisted 180 degrees counterclockwise (i.e.,
in a right-handed direction: most species of shell-bearing gastropods are still
right - handed, with notable exceptions, such as the Lightning-whelk (Busycon
contrarium Conrad, 1840). Many land and fresh-water species and even some
entire genera are also left-handed.) Internally, the digestive tract and nervous
system were twisted into a U-shape. The mantle cavity, gills and renal
and anal openings were now located in the anterior part of the body - i.e.,
just behind the head.
The problem of the large unmanageable pretorsion shell was solved with the evolution
of the asymmetrical coiling of the shell. Now, the coils were laid down
around a central axis (called a columella
and each coil lay beneath the preceding coil. In order to balance out
the weight of this shell, it shifted so that the axis of the spiral slanted
upwards and slightly backwards (the �asymmetrical � part!).
The shell was now positioned obliquely to the long axis of the body and the
gastropod could move about with relative ease.
(Large
Diagram)
Although there are fossil species showing the pretorsion plano-spiral shell,
all existing shell bearing gastropods have this post torsion, asymmetrical shell.
Several problems did arise for the gastropod as a result of this torsion however.
The main one being �fouling �. If the water circuit through
the mantle cavity had remained as it had been, the anus and nephridia
would have dumped directly on top of the head - a dangerous and not too nutritious
situation! Sanitation was thus a great a problem with this new shell design,
and the gastropods followed three different courses to solve this problem:
In one group, this problem was solved by the formation, over the mantle cavity,
of a cleft or split in the shell and mantle. At the same time, the anus
drew back from the edge of the mantle cavity and moved to a new position just
beneath the inner margin of this cleft. The inhalent current continued
to enter over the head and pass over the gills, but now instead of making a
U-turn, the water current flowed up and out through the cleft in the shell taking
with it the anal and nephridia wastes. Some of today �s gastropods
have retained this primitive cleft or shell slit.
Others modified their internal organ arrangement to have a single-gill arrangement
in a new mantle cavity, and developed an inhalant
water circuit to the side of the head. .Still another group underwent �detorsion
�, in which the twisting process was reversed and the mantle cavity and anus
once again opened posteriorly. A good example would be the common garden
slug.
Another consequence of this torsion and new shell position was that it restricted
the mantle cavity to one side of the body, and the opposite side of the body
was now pressed up against the shell. This compression resulted in a decrease
in size, or the complete loss of, the gill, auricle and kidney on that side
of the body.
So it came to be that about 500 million years ago, during the Cambrian period,
three basically different stocks, with different body-plans arose. Although
most of today's gastropods bear a single, asymmetrically coiled shell, some,
such as limpets and abalone have a flat saucer like shell. Still others
have no shell at all as is found in the sea and land slugs. These shell
changes and body adaptations resulted in the Gastropoda being divided into three
subclasses:
Taxonomy:
Proceed to Main Taxonomy
Table
- Class: Gastropoda
- Subclass: Eogastropoda
- SuperOrder:
Patellogastropoda
- Subclass:Orthogastropoda
The
article below this point has not been revised to match the above taxonomy (in
table) which is very much under construction.
Subclasses, etc. do not match up to the above table. The
taxonomy above is being based on the Academy of Natural Sciences the taxonomy
below is based on Brusca and Brusca's Invertebrates textbook.
1.
Subclass Prosobranchia: (proso-branch-i-a)
Latin: pros=front branch=gill
Greek: Proso-branchia
"Proso" = front, moving front and "Branchia" = Gills
(Branchio in single, Branchia in plural)
|
|
The
majority of the gastropods are prosobranchs. This group includes
all the gastropods that respire by means of gills and in which the mantle
cavity, gill and anus are located at the anterior of the body. They
possess a shell and torsion is evident.
Most prosobranchs are aquatic. Many have also developed a stone-like
(calcareous, made from calcium carbonate, like the shell) or horny (made
of chitinous material
similar to our fingernails, only not quite as hard) operculum. (For details
on this operculum, read
the foot and locomotion section)
Some members of this subclass are the limpets, periwinkles, conchs, whelks,
cones, murexes, cowries and volutes. Oddly enough, the most successful
family of all the molluscs, the Turridae, which are among the most
advanced Prosobranchs, are little known - they are often quite small,
and never found along the seashore. They have nearly 7,000 described
species so far.
|
| |
 Order
Archeogastropoda (or �Aspidobranchia �): Order
Archeogastropoda (or �Aspidobranchia �):
(Arch-e-o-gas-tro-poda)
Latin: arch=ancient eo=dawn gastro=stomach
poda=foot
Primitive prosobranchs in which there are two auricles, two kidneys and
two gills present. Nerve system is never concentrated. Shell
is either coiled or secondarily symmetrical as in the limpets. Largely
marine but there are a few that inhabit brackish water, freshwater or
even terrestrial habitats. Examples: Pleurotomariidae (Slit shells), Halitidae (Abalone), and Trochidae
(the Top shells). Most have operculi, and all are marine inhabitants.
|

Perotrochus
hirasei
(Pilsbry,1903) |

Haliotis rufescens
(Swainson, 1822)
Haliotidae
Red Abalone |

Tectus (Tectus) niloticus
(Linnaeus,
1758)
Commercial
Trochus orTop Shell
|
|

Janthina janthina
(Linnaeus, 1758) |

Crepidula fornicata
(Linnaeus, 1758) |

Stombus gigas
(Linnaeus, 1758) |
|
Order
Mesogastropoda ( or “Pectinibranchia”): (Me-so-gas-tro-poda)
Latin: : meso=middle gastro=stomac poda=foot
Greek: Meso-gatro-poda Meso =in the middle =pous=foot
Mesogastropods
possess one gill, one auricle and one kidney. An operculum may be present.
Mostly marine but a few do inhabit freshwater. This is the largest order
of gastropods and contains many common species, such as the Littorina,
Janthina, Crepidula, Stromb,
Lambis, Cerithium, Polinices, Vermetidae
worm-shelled snails, Pomatiasidae and Cyclophoridae.

Lambis lambis
(Linnaeus, 1758) |

Rhinoclavis vertagus
(Linnaeus, 1758) |
Polinices lacteus (Guilding, 1834) |

Vermicularia spirata
(Philippi, R.A., 1836 |

Pomatias elegans
(Mueller, 1774)
|

Alycaeus gibbosulos
(Stoliczka,1872)
|
|
|

Murex brandaris
& trunculus

Busycon carica
(Gmelin, 1791).
(Knobbed Whelk)
|
Order
Neogastropoda (Stenoglossa):
(Ne-o-gas-tro-poda)
Latin: neo=new gastro=stomach
poda=foot
Neogastropods possess a concentrated nervous system and usually a shell
with a siphonal canal. They are a carnivorous species having a radula
containing two or three large teeth in each row. Some possess a
poison gland. Nearly all have an operculum. All are marine inhabitants.
This group includes the beautiful Murex,
Busycon, Colus, Fasciolaria, Conus, Turridae
and Terebra.
|
|
|
| |
2.
Subclass Opisthobranchia: (o-pis-tho-branch-ia)
Latin: opistho=behind branch=gill
Greek:
Opistho=behind, back, Branchia=gills
|
|
The opisthobranchs display various stages of detorsion. Many have
adapted a secondary bilateral (i.e., two-sided, as in humans!) symmetry
in which the shell is either much reduced or completely absent. Gills
are generally posterior (i.e., behind) to the heart and are often on the
outside of their bodies in the form of plumes. They possess one
auricle (heart chamber) and one kidney. They are all marine inhabitants,
and many have adapted to a pelagic or swimming style of life. Most
are herbivorous, but many are parasitic (e.g. pyramidellas), living on
other bivalves and sea creatures.
Some of the more familiar opisthobranchs animals are; those with diminished
shells, the sea hares and those with no shell
at all, the nudibranchs and sea slugs. A few such as the bubble
shells do possess a hard shell.
|

Acteon eloiseae
(Abbott, 1973)
(Acteonidae / acteons)

Hydatina amplustre
(Linnaeus, 1758)
(Hydatinidae
/ Bubbles) |
Order
Tectibranchia:
Latin: tect=covered branch=gill
Shell
is present; however it is often much reduced or covered by the mantle.
They possess one true gill. Many are secondarily symmetrical.
All are marine inhabitants. Some members are the Acteon, Bulla,
Scaphander, Philine, Aplysia, Pleurobranchus, and the Pyrams.

Scaphander lignarius
(Linnaeus,
1758)
(Scaphandridae / Bubbles) |

Philine auriformis
(Suter, 1909)
(Philinidae) |
|
|
|
| |

Diacavolinia longirostris
(Blainville, HMD de, 1813)
(Cavollinidae / Sea butterflies) |
Order
Pteropoda:
Latin: pter=wing pod=foot
Greek: Ptero (Single) Ptrea (Plural), Greek not Latin
Here
the anterior portion of the foot has expanded to form swimming fins. These
sea butterflies �
(As opposed to the �sea hares �, which are different again!) may or may
not have a shell. The Pteropods are marine inhabitants and we know
such members as Spiratella, Clio, Cavolina, Limacina, and Cuvierina.
Their shells are most often found in fine, deep-sea sediments, where they
are not lost or crushed amongst coarser sand, gravel and rocks.
|

Polycera faeroensis
(Lemche, 1929) |
Order
Nudibranchia:
Latin: nud=naked branch=gill
These
are the shell-less sea slugs or nudibranchs. They are secondarily symmetrical (whatever
that means!). They do not possess a mantle cavity or gill.
Respiration is through the body surface, cerata or secondary gills located
around the anus. Their nervous system is concentrated. All
are marine inhabitants and we know them as the Doris, Dendronotus, Elysia,
and Aeolidia. As a bonus for us humans, they are amongst the most
beautiful creatures in the ocean - take a browse through the Nudibranch
and Sea Slug Sites found in the links
section, and I GUARANTEE you'll be surprised!
|
| |
3.
Subclass Pulmonata: (pul-mon-a-ta)
Latin: pulmo=lung (because they breathe air) |
|
The pulmonates retained the post torsion anterior position of the anus
and mantle cavity; however, the gills have disappeared and the mantle
cavity has become modified into a �lung �. They possess one
auricle and one kidney.
This subclass contains most of the woodland and garden snails. Garden
slugs are pulmonate snails that have evolved without developing a shell,
or that have perhaps lost them somewhere in the mists of time. Many
freshwater snails are also pulmonates.
|

Helix pomatia
(Linnaeus, 1758)
(Helicidae / Burgandy or Roman snail) |
Order
Stylommatophora:
Latin: styl=column omm=eye phor=carry
Greek: Stylo-mato-phora, Stylos = column, pillar Mato = Mati, Matia =
eye, -phora as prefix means "whom curry something"
eyes carried at the end of stalks (tentacle)
Stylommatophors
possess two pair of tentacles with eyes located at the tip of the posterior
pair. All are terrestrial. Included are the Helix
(the family of the famous French Escargot!), Polygyra, Pupa, Janella,
Deroceras, Philomycus, Palifer, Testacella and Limax.
|

Lymnaea stagnalis
(Linnaeus, 1758)
(Lymnaeidae / Great Pond Snail) |
Order
Basommatophora:
Latin: basi=bottom omm=eyes
phor=carry :
Greek: Baso- as prefix means, bottom or base, mati,matia,omata = eyes,
-phora
eyes carried at the base of stalk (tentacle)
Basommatophors
possess one pair of tentacles with eyes located near the tentacle base.
They are primarily freshwater inhabitants and they require air for respiration
although some do take water into their mantle cavity and have evolved
secondary gills. Some are terrestrial inhabitants and a few are marine
inhabitants. These include such familiar families as the Phasionellidae,
and many are left-handed (coiling to the left, instead of to the right,
like most marine species do). Most have thin, fragile shells, since
they don't have to put up with the rough-and-tumble of the waves in the
ocean.
|
|
Gastropod
Characteristics
|
(Generalized
Diagram)
The typical shell of the gastropod is the familiar conical spire
composed of tubular whorls. This shell, which is created, maintained, colored
and modified by the mantle, contains
the visceral mass of the animal: i.e., all its internal organs. Starting
at the apex, the smallest
and oldest part of the shell, whorls get successively larger and are coiled
about a central axis called the columella, which may be open or closed.
The largest whorl terminates at
the aperture or opening where the head and foot of the animal protrude.
(an
excellent diagram on the parts of a shell can be found on Peter Egerton 's Webpage
at: http://members.shaw.ca/bcshells/morphology.html)
A shell may be spiraled clockwise (right-handed shell) or anti clockwise (left-handed
shell). When a shell is held so that the apex (top) is up and the aperture
facing the person, those with the aperture facing to the right are right-handed
or dextral and those that open to the left are left-handed or sinistral.
Both sinistral and dextral shell can be found amongst members of
the some species. (Photo 1, Photo 2).
(An excellent article: "Reverse
Coiled Gastropods": by the Jackson
ville Shell Club on this subject)
The first shell whorls laid down by the larval gastropod (i.e., while it is
in its egg), are called the �protoconch �
(proto = before, conch is shell). It is represented by the smallest few
whorls at the apex of the shell, and is usually smooth, and lacks many of the
characteristics of the adult shell, often being colorless, or of a different
color from the rest of the shell.
A typical gastropod shell is composed of three layers; the outer periostracum,
the middle prismatic layer (Ostracum) and an inner nacerous
layer (Hypostracum). The periostracum is thin and composed of a horny
organic (made out of protein, actually!) material called conchiolin,
which is semi-transparent, being a brown color: the thicker the periostracum,
the darker the color, and the more the shell underneath is both protected from
sand grains and other abrasive elements of the animal's environment, as well
as from acidic water, which some of the hardier gastropods and bivalves can
survive in. Shell collectors often dissolve the periostracum of their shells,
so they can see the beautiful colors and patterns better. Scientists,
however, leave the periostracum on, since it is an important part of the shell.
The two inner layers are composed of calcium carbonate. In the middle
layer (which we normally think of as the outside of the shell, since it contains
the colors and patterns, the calcium carbonate is laid down as vertical crystals.
In the thin, inner, nacerous layer, the calcium carbonate is laid down in thin
horizontal sheet. Quite often, there are two or more sheets, each of which
reflect light differently, creating the shimmering effect called �iridescence �
(actually a product of refraction patterns - ask your physics teacher to explain!)
Reserve calcium carbonate is stored in certain cells of the digestive glad and
is used for shell repair or to add new growth thus enlarging the shell for the
growing animal. Molluscs can only form shells when they can extract CaCO3
(calcium carbonate) from the water, and keep it from being dissolved again.
Thus, if a lake or stream is acidic, or the soil is acidic (as in Coniferous
woodlands), shells, and therefore the animals that make them to protect and
support themselves, cannot survive. Also, below a certain depth in the
ocean (which varies with temperature and mineral content, calcium carbonate
cannot be deposited, since the water is under-saturated with Ca CO3. This
is called the �calcium carbonate compensation depth �, and no shell-bearing
molluscs can survive below this level. To summarize, shells can only be
formed in fresh waters that are non-acidic, and in the ocean at depths above
the level where the water becomes under saturated with Calcium Carbonate.
(A good site to see the
layers of a shell is at: http://members.lycos.co.uk/Mollusks/Schnecken/morphologie/schale.html)
Gastropods show an infinite variety of colours, patterns, shapes and sculpturing
of their shell (which is why people collect shells, as opposed to the entire
mollusc!!). In some gastropods, the shell is only conspicuously coiled in the
juvenile stages. The coiled nature disappears with growth, and the adult
shell represents a single large expanded whorl.
Examples of this are found amongst the abalones (Haliotis), limpets (several
families, including Lottiidae and Acmaeidae) and slipper shells
(the familiar Crepidula and Capulus). (The limpets became secondarily
symmetrical
during in their evolution.)
Haliotis rufescens
(Swainson, 1822)
Haliotidae
Red Abalone |

Collisella pallidula
(Gould, A.A., 1859)
Lottiidae
Snowy limpet |

Pectinodonta rhyssa
(Dall, W.H., 1919)
Acmaeidae |
Crepidula fornicata
(Linnaeus, 1758)
Calyptraeidae Common Atlantic slippersnail |
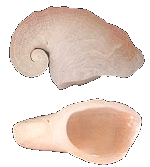
Capulus (Capulus) badius
(Dunker, R.W., 1882)
Capulidae |
|
In the family of Vermetidae (the
worm shells), the larval and juvenile shells are typical, but as the animal
grows older the whorls become completely separate. The adults look
much like a corkscrew, and sometimes don't coil at all, forming a tangled
mass of tubes called a �colony �
|
Vermicularia spirata
(Philippi, R.A., 1836)
|
Amongst many gastropods, the shell has become much reduced or is absent completely.
In other cases the foot and mantle are very large and the mantle has reflexed
backwards over the shell so that it becomes totally covered. These
animals are no longer able to pull their bodies completely into their shells.
|
The pulmonates show varying degrees of shell reduction and loss, culminating
in the slugs (land and sea), which have no shells at all.
|
Helix pomatia
(Linnaeus, 1758) |
The opisthobranchs have a much reduced shell which is closely related to the
degree of detorsion they have undergone. A few have well developed shells, such
as the bubbles, but most have a much-reduced shell that is often covered by
the mantle as is found in the sea hares.
Hydatina amplustre
(Linnaeus, 1758)
|
Polycera faeroensis
(Lemche, 1929) |

Aplysia parvula
(Guilding in Morch, 1863) |

Pyramidella dolabrata
(Linnaeus, 1758) |
Gastropods are able to withdraw into their shells by means of a retractor muscle.
This muscle, called the columellar muscle, arises from the foot and it is inserted
into the columella.
The most ancient gastropods, the abalones and limpets have two of these muscles
but in the more modern gastropods the left muscle has disappeared.
|
The typical foot of a gastropod is a large flat creeping sole similar
to the foot design of the ancestral mollusc. It has become adapted
for locomotion over a variety of surfaces.
|

Lymnaea stagnalis
(Linnaeus, 1758)
Great Pond Snail |
|
The limpets have become quite well adapted to clinging tenaciously to
the hard substratums (rocks, wood, other molluscs' shells, etc.) where
they live. Many marine and fresh water gastropods have adapted to living
on the soft sandy or muddy bottom. Others live on seaweed or terrestrial
vegetation or under rotting leaves and logs.
|

Cellana talcosa
(Gould, A.A., 1846)
Turtle Lmpet |
Typically, a pedal mucous gland opens onto the dorsal or ventral (i.e.,
top or bottom) surface of the foot. This secretes a slime trail over which
the animal glides. Waves of fine muscular contractions that sweep from
the anterior to the posterior (i.e., from the front to the back) of the foot
provide the power for locomotion.
The foot of many gastropods bears either a horny periostracum or calcium carbonate
disc, called the operculum.
This structure is found on the posterior dorsal (the back bottom) portion
of the foot. This is the operculum, and it acts as a �trap door �
that the animal can pull shut to close off the mouth of its shell, thus protecting
its soft body parts, which are safely inside. The operculum may also be
closed tight to guard against dehydration, if it should become necessary. (As
in during dry periods or winter (which in many parts of the world is just a
dry season), or when a pond dries up!)
( An excellent site to view operculums is: Websitium
Operculata)

Operculum (Cat's eye) from a Turbo petholatus |
|
|
Some gastropods, such as the marsh-dwelling pulmonates (Melampus,
for example, which can be found by looking at the high-tide mark of a
salt marsh, usually on or near the salt-marsh grasses, which are called
Spartina), extend the anterior portion of the foot and then pull
up the rest of their body behind it.
|

Melampus bidentatus
(Say, T., 1822 )
Eastern Melampus;Salt-Marsh Shell |
|
In another marine snail (Lacuna), the foot is divided into a right
and left half by a groove extending down the middle of their foot.
This snail moves by advancing one side of the foot then the other side
(a bit like walking!)
|

Lacuna vincta
(Montagu, G., 1803)
Banded chink shell |
|
The pelagic gastropods, the Heteropoda and the sea butterflies,
have adapted to a swimming life style. Being pelagic,
the foot has become modified into a powerful finlike, swimming apparatus
- in many species, almost looking like wings for flying in the water (hence
�sea butterflies �!)
|
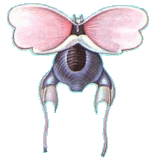
Diacavolinia longirostris
(Blainville, HMD de, 1813)
Long-snout Cavoline |
|
Some of the prosobranch have adapted for burrowing into the soft sand
or mud where they live. Here, the front of the foot called the propodium,
acts like a shovel. It has also developed a dorsal
flap-like fold of the foot that acts as a protective shield for the head.
This mode of living is more common in the Pelecypoda (bivalves), however.
|
An example
is the Olivella shells
See: Jaxshells
|
A few gastropods are sessile (i.e., once they settle down somewhere, they don't
move at all (remind you of any couch potatoes you know??)). They usually
attach themselves to the shells of other living or dead molluscs.
The foot has adapted to become a sucker-like. The Worm shells
are totally immobile and are either attached to other molluscs or entangled
in sponges.
| Water
Circulation & Respiration: |
Gastropods have developed many methods of attaining oxygen from
the water or land habitats where they live. To some extent the exposed
body surface, especially that of the mantle, plays a varying role in the respiration
of all the gastropods. Most breathe by means of a gill/s (ctenidia) or
secondary gill structures.
|
In the prosobranchs with cleft shells (slit shell,
Scissurellidae (which are like mini-slit shells!), and Fissurellidae
(the Key-hole limpets) the most primitive type of gill structure and water
circulation occurs. In these gastropods there are two primitive
gills and the rectum and anus open beneath the shell perforation or cleft,
some distance away from the mouth.
|
Perotrochus
hirasei
(Pilsbry,1903)
|
|
In the abalone (Haliotis), the shell contains a row of perforations.
The mantle is split along this line of holes. Inhalent
water is pulled into the anterior portion of the body by the action of
the lateral cilia (tiny hairs) on the gills. The outstretched gills
divide the mantle cavity into a ventral
inhalent chamber and a dorsal
exhalent chamber. Water flows into the inhalent chamber, passes
through the gills, then into the exhalent chamber and finally exits through
the shell perforations - an ingenious arrangement, actually!
|
Haliotis rufescens
(Swainson, 1822) |
|
The Scissurellidae have a similar system but instead of having
shell perforations, the exhalent current
passes out through the long, narrow notch at the posterior mantle edge
where the anus exits.
|

Scissurella coronata Watson, 1886 |
|
Keyhole limpets (Fissurellidae) have conical shells that either
have a hole at the apex or a cleft at the anterior margin (i.e., the front
end). The mantle
extends through this opening or slit, forming a siphon through which the
inhalent water is sucked. This inhalent water then passes over the
gill and
|

Diodora aspera
(Rathke 1833)
Rough Keyhole Limpet |
|
exits
at the posterior edge of the shell opening (so, it is sucked in the top
or the front, through a hole of slit, passes over the gills, then exits
out the back - it just sounds so much more scientific when said the other
way? (NOTE: Actually scientists don't use technical jargon just to impress
or confuse non-scientists (although it sometimes seems that way!!), but
because each branch of science has developed a more exact, or precise
vocabulary than we use in everyday speaking: the work �anterior �,
for example, has only one meaning, which is precisely defined and
cannot be confused with any other word - the word �front �,
on the other hand, could either mean the head area, or the part of the
animal that is going forward - which might not always be the head area!
So, it is better in a scientific context or situation, to use the word
�anterior �: than �front �, because it only has
one meaning, while front has at least two! Half the trick to science
is learning how to translate from jargon or �science-speak �,
to ordinary language - hardly anyone ever truly THINKS in jargoneese!!)
|
|
True limpets lack the hole or cleft and the mantle have developed an overhang.
This overhang forms a pallial groove on each side of the foot. The
inhalent water enters from the anterior of the shell, splits into two
streams flowing into these two grooves. It then passes through the
gills merges into a single stream at the posterior and exits out as a
single exhalent stream.
|

Acmaea filosa
(Carpenter, 1865) |
The remaining prosobranchs have undergone major gill structure and water circulation
modifications. The entire gill axis is attached to the inner, or body
side, of the mantle cavity. They have only a single left gill with filaments
that are formed on one side of the axis. Water enters the mantle cavity
to the left of the head and exits on the right side. To prevent fouling
of the inhalent water, the rectum has become elongated and the anus exits near
the right mantle edge in the region of the exhalent water current.
|
Many prosobranchs have improved on this system by developing a spout-like
inhalent siphon formed by a folding of the mantle �s edges. Some
gastropods even carry this a step further and the anterior edge of the
shell has evolved to become a grooved, elongated extension to house this
siphonal canal. (A good example of this is found amongst the Tibia
shells that have carried this to an extreme in some instances.)
|

Tibia fusus
(Linnaeus, 1758).
Shinbone tibia |
Some prosobranchs have left the water environment entirely. These are
the operculate land snails, and are not true pulmonates. They have evolved
a �lung � from the mantle cavity, as have the pulmonates. Most
of these prosobranchs are restricted to living in moist tropical environments
in order to maintain their fluid balance.
In the opisthobranchs where partial detorsion (untwisting) has occurred, there
has been a loss of the original gill structure and a secondary gill has evolved.
|
In
the nudibranchs and sea slugs where complete detorsion
has occurred, the mantle cavity and gill have disappeared all together.
Respiration takes place through the general body surface or through a
secondary gill (i.e., a structure which is not really a true gill, but
which performs the same function.). To help
|

Dirona albolineata
(MacFarland in Cockerell & Eliot, 1905)
Alabaster dirona |
|
increase
the body surface for this absorption, some have developed numerous projections
called cerata. These cerata are usually arranged in rows.
Not all opisthobranchs have these cerata however. Their cerata often contain
such brilliant colours as red, yellow, orange blue and green, which makes
them incredibly beautiful!. Some slugs are smooth while others have developed
secondary gills arranged in a circle around the anus. Sea slugs
and nudibranchs are amongst the most attractive molluscs one could ever
see.
|
|
In the Pulmonates, the gill has disappeared totally and the mantle
cavity has modified to become a highly vascularized (i.e., it has lots
of blood vessels to absorb O2 and give up CO2) primitive lung. The
mantle edges fit tight against the foot, except for a small opening on
the right side called the pneumostome. Respiration occurs when
mantle floor (acting like a diaphragm) tightens and flattens. This
causes the mantle cavity volume to increase, which in turn sucks air in
through the pneumostome. As the air pressure increases the pneumostome
closes, to hold the air in. The muscles of the mantle then relax
and arch upwards which increases the gas pressures in this cavity. This
increased
|

Pomacea bridgesii (effusa)
For an excellent photo of the pulmonate gill of an apple snail, Please
vistit the AppleSnail site |
|
pressure facilitates (i.e. assists or makes easier) the absorption of
oxygen into the highly vascularized mantle wall chambers and also causes
the air to be forced out.
|
Many fresh-water snails are actually
terrestrial Pulmonates that have returned to the aquatic environment. Some
must return to the water surface to breathe, while others have developed a long
retractable siphon from their mantle that they use much like a snorkel (!).
Others can absorb oxygen directly through the mantle surface from the water
they draw into their mantle cavity. In still another group, the mantle
cavity became much reduced and a conical projection from the foot called the
pseudobranch developed as a secondary gill.
The gastropods have an open circulatory system - the basic circulatory system
found in most molluscs. They have a hemocoel, or open cavity, into which
the blood (called �hemolymph �, for some reason) is pumped. Oxygenated
hemolymph is collected from the gills or mantle cavity and pumped into a number
of open sinuses. Here the tissues and organs are literally bathed in this
oxygen-rich blood. As it passes over the tissues in these sinuses, it
flows into the ctendial (gill) vessels where gas exchange takes place.
It is then again drawn into the atria to be pumped out of the heart. (Diagram)
|
The gastropod heart is located anteriorly (i.e., closer to the head) in
the visceral mass. In all but the primitive archaeogastropoda, the
right auricle has disappeared or become vestigial due to the loss of the
right gill - one of the results of torsion, as discussed above.
The ventricle gives rise to a single, short aorta, which then branches
posteriorly to provide the visceral mass with blood, and anteriorly to
supply the head and foot. An enlargement in the anterior vessel
- a sort of �second heart �, functions in controlling blood
pressure. Blood from the kidneys usually enters the brachial circulation,
but in some cases it returns
|

Architectonica perspectiva
Linnaeus, C., 1758) Clear/Painted Sundial
(an archaeogastropoda) |
|
directly to the heart. In a few Gastropods, such as the family Planorbidae
(which you aren't likely to encounter, by the way), the plasma contains
hemoglobin instead of hemocyanin. (Hemoglobin uses Iron for transporting
Oxygen and Carbon dioxide, while hemocyanin uses copper. Thus, the
blood of most molluscs is a light greenish-blue, instead of the red we
usually associate with blood, which comes from the iron in hemoglobin).
|
In gastropods, the nervous system is distinctly ganglionated (i.e., it
has well-defined and specialized nerve cells) and is somewhat complex.
It is quite asymmetrical, and twisted into a figure eight as a result of torsion.
A pair of cerebral ganglia (which function as a small brain) give rise to nerves
anteriorly that connect to the eyes, tentacle and a pair of buccal ganglia.
The buccal ganglia innervate
(send nerves to) the muscles of the radula and adjacent structures.
A nerve cord extends ventrally from the cerebral ganglion (located on each
side of the esophagus) and gives rise to the two pedal nerve cords. These
two cords extend to the midline of the foot to another pair of ganglion, which
in turn innervates the foot muscles.
Another pair of nerve cords issue from the cerebral ganglia also. These
are the visceral (the viscera are the body organs) nerves, and they travel posteriorly
(i.e., away from the head) until they finally meet in a pair of visceral ganglia.
Between the cerebral ganglia and the visceral ganglia and along the visceral
nerve cords lay two more sets of ganglia
(NOTE: so, instead of having one large, complicated brain like we do, gastropods
have 8 tiny, very simple brains, which coordinate between themselves, although
each pair have specialized functions.) Firstly are the pleural ganglia,
which innervate the columellar muscle and the mantle. The pleural and
pedal (pedal means foot. remember) ganglia are then joined together by
means of a pair of connective nerve fibers. Secondly and more posteriorly
are the parietal ganglia. These innervate the gills, osphridia,
and the mantle. They send out nerve fibers to the various structures of
the viscera as well).
All gastropods display some degree of ganglial concentration. For example the
pleural and cerebral ganglia are always adjacent to each other. In many
cases the visceral ganglia have been fused together to form a single nerve center.
| In the genus Haliotis
(Abalones), the pedal and pleural ganglia have become fused, and send a
long pedal nerve to the foot. In the Busycon (which include the famous
left-handed Lightning Whelk), all except the visceral ganglia have migrated
forward and are located around the esophagus and just below the cerebral
ganglia. Here all ganglia connectives have been lost except those
between the parietal and visceral ganglia. |
 Haliotis asinina
Haliotis asinina
(Linnaeus, 1758)
Donkey Ear Abalone
|
|
In the pulmonates, even the visceral ganglia have migrated forward, which
has resulted in a secondary bilateral symmetry of the nervous system.
|

Planorbis planorbis
(Linnaeus 1758)
Ramshorn Snail |
|
Opisthobranchs that have undergone complete detorsion, possess nervous
systems that have become symmetrical again, in simple bilateral fashion.
|
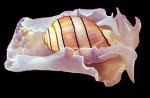
Hydatina amplustre
(Linnaeus, 1758)
|
Gastropods possess the following sense organs: eyes, tentacles, osphridia
(olfactory organs), and statocysts (organs of equilibrium, like our inner
ear: they help the mollusc tell which direction is �up � - not always
easy in water when you aren't on the bottom, or sometimes even when you are).
|
- Eyes:
Most gastropods have eyes located at the base of cephalic tentacles. Most are very simple
open pits containing only photoreceptor (photo means light) and
pigment cells. In the more advanced gastropods the pit has closed
over and evolved to contain a proper cornea and lens. The most highly
developed eyes of all the gastropods are found in the pelagic sea hares,
however most gastropoda eyes are but simple light detection organs.
(Diagram)
|
|
-
Tentacles: Prosobranchs possess a single pair
of cephalic (on the head, i.e.) tentacles. Pulmonates and Opisthobranchs
have two pair. These tentacles, in addition to bearing the eyes, contain
tactile (i.e., touch) and chemoreceptor cells. In pulmonates the second
pair of tentacles are knobbed. In nudibranchs, the distal (more distant,
as opposed to proximal, or closer) half of the tentacle wall displays plate-like
folds called �rhinopores �. These knobs and folds increase
the surface area for chemoreception.
-
Osphridia:
The evolution of the osphradium
(See note) closely parallels that of the gills. An osphradium is available
for each gill present. The osphradium has become either filamentous
or folded to increase its surface area. The leading theory amongst
scientists is that the function of the osphradium is to detect sediments
in the water passing over their gills, but nobody really knows for certain!!
(NOTE: Latin is tricky, but in science one
has to put up with it, since many scientific terms are from Latin, which
was once-upon-a-time the language scientists all over the Western World
(Europe and the Middle East) used to communicate. English is now taking
that role, although it has not become as fully universal as Latin once was.).
- Statocysts:
These organs of balance are generally located in the foot near the pedal ganglia,
however, some of the opisthobranchs (especially the nudibranchs), these have
migrated forward to a position next to the cerebral ganglia
| Nutrition
& Digestive System: (Diagram) |
Gastropods exhibit virtually every type of feeding possible. Members
of this class can be herbivores, carnivores, scavengers, ciliary feeders, or
parasites. Despite these feeding differences, some generalizations can
be made:
-
Gastropods
almost always employ a radula (Diagram) in feeding and in
many cases this has become a highly developed feeding organ. The radula
may contain as few as 16 teeth (with only a couple of them fully functional,
as in Cones and Turridae, which use them as poison-tipped �spears �.)
Or as many as 750,000 teeth arranged in rows. The action of this radula
may be that of a grater, a rasp, a brush or a comb, or spear.
-
Digestion
is always at least partially extracellular in gastropods - there are no
gastropod species where all digestion takes place inside the individual
cells.
-
The
enzymes needed for extracellular digestion are usually produced by the salivary
glands, esophageal pouches, and/or the digestive gland, or by a combination
of all of these structures. There are few exceptions to this norm.
-
The
stomach is the site of extracellular digestion and the liver is where absorption
and of intracellular digestion takes place, in species where this occurs.
-
Food
is passed through the gastropod gut, or at least partly, by the action of
the ciliary tracts.
-
As a result of torsion, the stomach has been rotated 180 degrees so that
the esophagus enters the stomach posteriorly and the intestine leaves it
anteriorly (the opposite of most animals!). In some of the higher
Gastropods, the esophagus has started to migrate forward again.
- The enzymes
needed for extracellular digestion are usually produced by a) the salivary
glands, b) esophageal pouches, and/or c) the digestive gland, or by a
combination of all of these structures.
The most primitive digestive system is found in the Archeogastropoda: Keyhole
limpets feed primarily on sponges that are rasped from the substratum by the
radula. The salivary glands in these limpets only secrete mucus, which is used
for radular lubrication and food transport. The esophageal pouches are
well developed and they as well as the digestive gland produce the enzymes necessary
for extracellular digestion.
The
area of the stomach nearest the esophagus
is partially lined with tough chitin and
contains a ridged sorting region. The end nearest the intestinal opening
is conical and forms a style sac. A deep groove runs the length of the
stomach. The sorting area directs food particles towards the style, where
some of these particles pass through the style sac groove. The digestible material
is directed and passed into the ducts of the digestive gland. The rest is compacted
and passed directly into the intestine. Digestion is primarily extracellular.
(Diagram)
Most gastropods are herbivores, carnivores or scavengers. They have lost
this primitive chitinous lining, sorting area and style sac of the ancestral
digestive system. Digestion has become totally extracellular.
-
The
Herbivores: Many prosobranchs, opisthobranchs
and most Pulmonates are herbivores. The radula in these gastropods
generally contain numerous small teeth. The upper margin of the buccal
(i.e., mouth) cavity bears a chitinous jaw (in the pulmonates a lateral
pair of jaws may also be present). Often, the esophagus is enlarged
to form a crop (food-storage area) just before the stomach opening.
Here food is temporally stored and enzymes from the salivary and digestive
glands begin to break down the food particles.
- The Carnivores:
Most carnivorous gastropods are from the prosobranchs and opisthobranchs
subclasses, however a few are also pulmonates. In this group, the radula
contains fewer but larger teeth. In most of the prosobranchs, the jaw
has disappeared -the buccal cavity has become folded to produce an extendable
proboscis and proboscis cavity, that opens to the outside through the mouth.
When feeding, this proboscis can be extended from the mouth a considerable
amount. At the tip of the proboscis is the radula and food enters the proboscis
cavity instead of the buccal cavity. The radula is retracted when feeding
is finished.
In some prosobranchs, such as the Muricidae,
Buccinidae (the �true � whelks (example: the Neptunea
shell, found on both the Pacific and Atlantic coasts of N. America)
and Naticidae, the radula is adapted for drilling neat circular
holes in the shell of its host. Secretions coming from an eversible
gland (i.e., it can be extended out of the body, like the stomach of a starfish
or sea urchin) located on their foot aids this drilling. Though not
fully understood, these secretions may be a form an acid that dissolves
the prey's shell.

Chicoreus ramosus
(Linnaeus, C., 1758) |

Neptunea lyrata
(Martyn, T., 1784) |
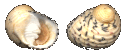
Nerita versicolor
(Gmelin, 1791) |
The posterior part of the esophagus of some carnivores is enlarged.
This enlargement may act as a crop
or a gizzard. When this structure is present, the prey is swallowed
whole and then the chitinous plates lining the gizzard walls crush it (ouch
- what a way to go!!)
Cone shells are very remarkable
amongst the carnivores. The cones produce very toxic venom in a poison
gland that opens into the buccal cavity. The radular teeth, which are
attached to the radular membrane by a slender cord of tissue, have evolved
to become sharp, hollow, barbed tubes which contain this venom: a bit like
a poison-containing spear! The proboscis of cones is highly maneuverable
and can be shot out with explosive force and speed. When the proboscis
is projected, a single radular tooth slips out of the radular sac into the
buccal cavity and the proboscis strikes. The tooth is rammed into the
victim much like a harpoon. The tooth breaks, releasing its poisonous
contents into the victim and the venom quickly immobilizes it. Cones
feed mainly on fish, other molluscs and annelid worms. For an AWESOME
little movie of a cone-shell actually harpooning and eating a fish, go to:
http://grimwade.biochem.unimelb.edu.au/cone/envenom.html.(NOTE:
It is a 2.2 meg file, so beware - download takes 20 min. with a 28.8 modem
(a few seconds with a t-1 or lan, however!). The cone venom acts quickly,
paralyzing the nerve-muscle junction of its prey. Some of these cone
neuro toxins can be very toxic or even fatal to man. Death can result
in a matter of hours. On the other hand, man has discovered ways of
using these poisons to develop potentially life saving drugs. (Link to Man
and Molluscs article, the section
on Medicine)
The digestive systems of many carnivorous nudibranchs have developed several
modifications. Some have small blade-like jaws in their buccal cavity
that are used to cut pieces of tissue from their prey. They don't possess
a proboscis, or esophageal pouches: the food passes into a simple ciliated
stomach. These cilia sweep the food particles posteriorly towards the
3 to 5 ducts of the digestive gland. In the Nudibranchs, these digestive
glands are confined entirely to their strange cerata:
Each cerata contains one tubule that is then joined together with the other
tubules in a series of branching ducts leaving the stomach. The secretory
cells of these digestive tubules pour out digestive enzymes. Absorption
takes place extracellulary in the stomach walls and digestive gland.
The stinging cells of some of their prey are engulfed (but not digested) in
the cerata of the nudibranchs, and are harmlessly moved into the distal tips
of the cerata (called cnidosacs), which open to the surface. These �nematocysts
� are then used for the nudibranchs � protection!! (Diagram)
- The Style Bearers
and Ciliary Feeders: Crystalline
styles are found in many ciliary feeders. A crystalline style is a firm
gelatinous protein (containing carbon-splitting enzymes) rod that lies
in the stomach at the intestinal end. It is embedded in a ciliated style
sac with its tip projecting into the stomach. When these cilia beat,
the style begins to rotate and the projecting end rubs against the chitinous,
plate-like, gastric shield found on the stomach wall. This abrasive
action erodes the style, releasing the enzymes into the stomach. The
rotation of the style does double-duty, also aiding in the mixing of the stomach
contents. (Diagram)
Some
gastropods live entirely on phytoplankton or organic detritus. Their
shell and mantle edges are held tightly against the substratum except for
a small gap on each side of the anterior (i.e., forward-facing) region.
Water containing miniscule food particles enters the left side of the mantle
cavity. Here these particles are trapped in mucus, which covers the
long gill filaments. The water then passes out the right side.
The trapped food particles pass into a longitudinal groove lying along the
side of the body and in contact with the gill filaments. Here the
food particles and mucous are compacted into a mucous string. The
anterior potion of the groove is located near the mouth. Here the
radula seizes a section of this string and pulls it into the buccal cavity
at regular intervals. These food strands are then passed onto the
stomach through the esophageal food groove. Here the crystalline style
discussed above, assists with the digestive process.
Products
of digestion and other waste materials are then passed into the digestive
gland where digestion completed intracellarly and absorption occurs.
A deep groove parallel to the style carries the waste particles to the intestine
for excretion.
Some
gastropods such as the family Strombidae [Lambis (the �sea
spiders) � and Strombus (the �true � conchs)] are
style feeders as well. In these molluscs, algae is scrapped off the
substratum (i.e., whatever they live on) by very delicate radular teeth.
The Strombus produce amylase and cellulase, which assist the style
in digestion.
|
|
|
|
Lambis
lambis
(Linnaeus, 1758)
|
Stombus
gigas
(Linnaeus, 1758)
|
Another
ciliary feeder is the shell-bearing sea butterfly, the Limacina (See:
Limacina helicina).
This gastropod has no crystalline style, although a vestigial sac is present.
They feed primarily on diatoms and Foraminifer (Good Site: Foram
Gallery to learn more about Foras) Their feeding methods are complex
and fascinating: the mucous-coated walls of the mantle cavity trap the minute
food particles when they are pulled through with the inhalent water current.
It is then compacted into mucus strings and a ciliated tract carries it
to the buccal cavity where radula pulls off sections. The posterior
esophagus is extended and acts as a pump as well as a gizzard. The
walls in this sac contain four large opposing teeth that fit closely together
and they crunch the diatoms and Foraminifer shells. This action also
tends to move the food along the esophagus and into the stomach. Enzymes
are produced by the digestive gland and absorption is entirely extracellular
and takes place in the stomach.
- The Parasites:
A parasitic lifestyle has developed in a few of the gastropods. The
most noticeable example of this is to be found in the Pyramidellacea,
which live as ecto-parasites (living on the outside) of Polychaetes
(a multitudinous class of marine worms) and other marine bivalves. The
proboscis here is equipped with a sucker and stylet for gripping and piercing
the host. The tissue fluids are then ingested by a pumping action of
the buccal cavity.
(See: "Parasitic
Mollusca", article by Felix Lorenz )
There
are many groups of ectoparasitic snails on starfish and Sea-urchins. Most
of these belong to the Opisthobranchia. Thes opisthobranchia have retained
their shell. Only the very tip of this shell protrudes from their
host �s exterior surface. (See: Opisthobranchs
from the Mediterranean Sea and elsewhere )
The
Entoconcha, a small worm-like gastropod, lives in the gonad of
the sea cucumber, Synapta. By attaching their mouth to one
of the host �s blood vessels, they are able to feed upon its blood.
(Sounds like Sea Vampires!)
| Excretion
& Water Balance: |
As a result of torsion, all members of the gastropoda class except the Archeogastropoda
(they still have both nephridia)
have lost the right nephridia, or it has just been partially retained as a part
of the reproductive duct. The nephridium is a U-shaped sac and the walls have
been greatly folded to increase the surface area for secretion. It is
located anteriorly in the visceral mass.
Excretion involves filtration onto the coelom and reabsorption and secretion
in the nephridium. The nephridium in most cases drains into the pericardial
(peri =around, cardial=related to the heart, hence �around the heart �)
cavity via a small reno-pericardial canal and the wastes are excreted through
a short ureter (nephridiopore).
In opisthobranchs and prosobranchs,
the nephridiopore opens at the back of the mantle cavity and the wastes are
carried away with the exhalent water. This cannot happen in the pulmonates
however, as the mantle is working as a lung. The ureter has lengthened along
the right wall of the mantle and it opens on the outside, near the anus.
Aquatic gastropods, like most aquatic invertebrates, excrete ammonia or ammonia
compounds. Terrestrial pulmonates convert this ammonia into relatively
insoluble uric acid and water. This adaptation helps them to conserve
their valuable body moisture. However, they still do lose valuable moisture
to the air through their body surface and as a result many can only survive
in moist tropical climes of the world. Some, such as the Annulariidae,
have developed a small shell-like tube and use this to obtain air when their
operculum is closed.
Others have become nocturnal to avoid the heat of the day, or they live beneath
moist, decomposing vegetation. During hot dry periods or in the colder
months in the temperate regions, they burrow into humus or soil and become inactive
(this is called �estivation �,
which is like an extreme form of hibernation, except that metabolic rates approach
zero. This means that molluscs in this state can survive many years waiting
for favorable conditions to revive them!). They draw the edges of their
mantle together and they then secrete a thin protective calcareous membrane
in front of their shell aperture. Fresh water snails estivate when ponds
dry up and hibernate when they freeze over.
Studies have shown that the digestive gland of most gastropods also plays a
roll in the excretion of wastes. Excretory cells in this gland empty out
into the stomach and intestine.
The Gastropods are a mixture of dioecious
(two sexes) and hermaphroditic
(one sex) groups. Most possess either a single ovary or single testis
located in the spirals of the visceral mass next to the digestive gland.
Often, elaborate courtship rituals proceed the actual mating.
In the more ancient gastropods, the Archeogastropoda, the gametes pass through
short ducts and into the right kidney then they pass into the mantle cavity
via the nephridiopore. Eggs are provided with, at the most, a simple gelatinous
envelope, which is produced by the ovary. There is no need for copulatory
organs as fertilization takes place in the open water after the eggs and sperm
have left the mantle cavity.
In all the other gastropods the right nephridium has degenerated except for
the portion that functions as part of the genital duct. The genital duct
becomes considerably longer and undergoes differentiation to provide for sperm
storage and egg membrane formation. This longer duct (pallial) leads directly
to an opening in the mantle cavity.
In gastropods where the reproductive system provides for tertiary (i.e.,
tough external membranes which protect the egg better) egg membranes, the males
have had to develop a penis so that fertilization can take place prior to membrane
formation. This penis is a long extension or fold of the body wall just
behind the right cephalic tentacle. The entire male duct consists of a
coiled duct from the testis (pl. testes - gastropods have only one.), a short
renal portion to the vas deferens and the pallial vas deferens, containing the
prostate.
Sperm from the penis is transferred to the female where it is then stored in
the end of the pallial oviduct, where the eggs are fertilized. In the
female reproductive system, the pallial section of the oviduct has modified
to form an albumin (egg white) gland and a large jelly or capsule gland.
Eggs are either formed in jelly masses or are enclosed in a gelatinous capsule
after fertilization. These eggs then pass through the oviduct to the outside,
where they are on their own!
A few prosobranchs, all opisthobranchs, and pulmonates are
both male and female (hermaphrodites). The single gonad produces both the sperm
and the eggs. The genital duct becomes divided into two channels for the passage
of both sperm and egg. Two meeting hermaphrodites often go through complex
courtship rituals before the mutual exchange of sperm via penis in oviduct.
Fertilization is reciprocal.
Some gastropods, such as the slipper shells (e.g. Crepidula fornicata
L., 1758) that live stacked on top of each other, start life as males.
Then the male reproductive tract degenerates and the animal then either develops
a female reproductive system or another male system. An older male will
remain a male if it is attached to a female. The presence of a large number
of males will influence certain males to become females. Once female,
however, they remain as females forever. (In the world of Crepidula,
feminists RULE!).






